
| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
We began by addressing the difficulties with accessing MasteringBiology and by the Reading Quiz closing before the actual deadline. Students who have obtained an access code can email Kelly Howe ( khowe@unm.edu) to get an extension of the Reading Quiz.
I will do everything possible to allow you to succeed in this course, but you have to do your part:
A First Look at Your Own Genome. In the last part of the course, we will be discussing personal genomics. If you are interested in having a first look at your own genome at no cost to you, submit an essay of no more than 1,000 words on what you think you would learn from the experience (to pszauter@unm.edu). The writer of the best essay will receive a free genome analysis from 23andMe. The deadline for essays is midnight on Friday, September 13, 2013.
This is not a class assignment, and no extra credit will be awarded for participation. You will not be required to share any of the results of the analysis of your genome with anyone, ever. You can read about one person's experience at OpenHumanGenome.
In the last lecture, we reviewed ten key discoveries in genetics that built the science. The first was Mendel's work that established the particulate nature of the gene. This work was widely distributed but poorly understood for over 35 years until the rediscovery in 1900. What made the work more widely comprehensible was Flemming's description of chromosome behavior during mitosis, which we discussed last time. There was a period of time during which chromosome behavior in mitosis was understood, but there was no description of meiosis until Sutton and Boveri described it independently in 1903. During this interval, it was not clear how sexual reproduction could occur without doubling the chromosome number every generation.
We reviewed the basics of mitosis and meiosis from last lecture, noting that the purpose of mitosis is to make identical cells, while the purpose of meiosis is to produce haploid cells that will become gametes. This is accomplished through two cell divisions. The first division of meiosis is a called the reductional division because it reduces the number of chromosomes from the diploid number to the haploid number. The second meitotic division resembles a mitotic division of haploid cells.
We briefly reviewed the central role of meiotic recombination (crossing over) in creating chiasmata, which together with sister chromatid cohesion, hold together the bivalent or tetrad so that it can achieve proper orientation at metaphase I (see the figure). Once all kinetochores are oriented to an appropriate pole, the kinetochores no longer send an inhibitory chemical signal and the Anaphase Promoting Complex activates the protease separase to cleave the cohesin complexes that maintain sister chromatid cohesion, initiating anaphase. We used slides and physical models to illustrate this.
We reviewed three aneuploid syndromes from last time:
Trisomy 21 (Down Syndrome): Individuals with an extra copy of chromsoome 21. This produces mental retardation, although recently there have been great strides in raising these individuals in an enriched environment to improve their development. They commonly have heart defects, usually have some extent of hearing loss, and are at greatly elevated risk of leukemia. They have a characteristic facial appearance that is easily recognized.
Turner Syndrome (45,X): Individuals monosomic for the X chromosome and lacking a Y chromosome are said to have Turner Syndrome. They are phenotypically female and are recognized at birth by a webbed neck, which is easily corrected surgically. They show primary amenorrhea (they never begin to menstruate) and are sterile. They have short stature and a broad chest. They have specific cognitive defects but are not considered retarded.
Klinefelter Syndrome (47,XXY): XXY humans are phenotypically male. They are not always recognized at birth, but the condition is normally apparent at puberty. They have hypogonadism and reduced fertility. They have less muscle mass than is typical for males and have a female-like distribution of body fat, with broad hips and gynecomastia. They are at increased risk for diseases that have a higher prevalence in females, like autoimmune disease and breast cancer. They have some specific cognitive defects, but are not considered retarded.
We would like to understand the origin of these aneuploid syndromes. This would mean being able to identify at which meiotic division nondisjunction occurred, and in which parent, to produce the aneuploid offspring.
We would also like to see if the aneuploid syndromes can tell us anything about the role that chromosomes play in development.
Turning to the first problem, we reviewed the consequences of nondisjunction at the first meitoic division and at the second meitotic division, first using a simplified set of drawings with no crossovers, then using a more complicated set with crossovers. Consider a set of disomic gametes resulting from meitotic nondisjunction. It is clear using drawings and models that nondisjunction at the first (reductional) division will result in disomic gametes that have both homologous chromosomes, while nondisjunction at the second (equational) meitoic division will result in disomic gametes that have sister chromatids. When we add exchanges, this scrambles the results a bit until we see that we can tell the division at which nondisjunction occurred by following the centromere. Disomic gametes that have nonsister centromeres arose from nondisjunction at the first division, while disomic gametes that have sister centromeres arose from nondisjunction at the second division. This is easy to remember when we note that the function of the first (reductional) division is to reduce the chromosome number to the haploid number; if nondisjunction occurs at this division, the number of homologous centromeres is not reduced, and the disomic gametes will contain nonsister centromeres. The summary diagram is shown below.

A student pointed out that in the drawing above, the disomic gametes with crossovers do not contain two chromosomes that are reciprocal products. This is correct, and it is deliberate. The chromosome models used in class are three-stand double crossovers. One chromatid has two crossovers, its sister chromatid has no crossovers, and the other two chromatids have one crossover each. It is also possible to have four-strand double crossovers (each chromatid has one crossover) or two-strand double crossovers (two chromatids have no crossovers and two chromatids have two crossovers). It is left to the student to draw out these possibilities and to consider why all of these possible crossover types are equally effective at creating chiasmata.
If we accept that there is a large amount of genetic variation in human beings that can be detected by unspecified molecular techniques, we can see in principle that it would be possible to examine the DNA of a person with Down Syndrome (and their parents) to determine from which parent the extra chromosome was derived, and whether the nondisjunction event that gave rise to the disomic gamete occurred at the first meiotic division or the second meitoic division. This requires that at some level, both the maternally-derived and paternally-derived chromosome in the genome of each parent can be distinguished. In response to a question, I had to point out that every human being has both a maternally-derived and a paternally-derived chromosome set. From the standpoint of his or her offspring, however, all chromosomes derived from his or her father are the paternal set for that individual, regardless of whether they are ultimately derived from the paternal grandmother, the paternal grandfather, or as usually the case, the paternally-derived set is a combination of genetic material from both paternal grandparents.
An analysis of a large number of such cases, in which the individual with Down Syndrome and both of his or her parents are examined using molecular techniques, produces the following conclusions:
Please see the OMIM entry on Down Syndrome for more detailed information and references.
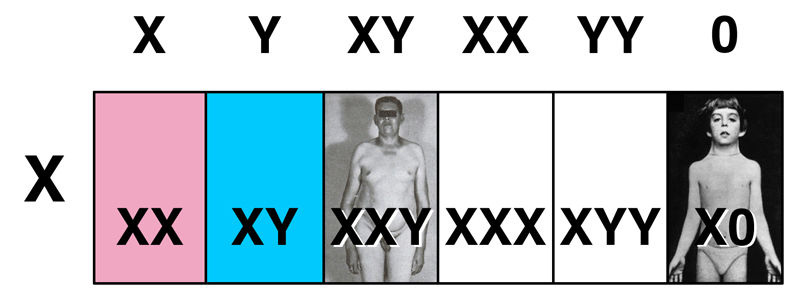
We turn our attention on the sex chromosome aneuploid syndromes, Turner Syndrome (45,X) and Klinefelter Syndrome (47,XXY). How do these arise?
First, consider the case of normal meiosis in both the mother and the father. Females produce eggs with a single X chromosome, while males produce sperm with either an X chromosome or a Y chromosome. We can predict the offspring as shown in the Punnett square below.

In this case we see that normal meiosis in both parents produces an equal number of normal male (XY) and normal female (XX) offspring.
Consider the case in which maternal nondisjunction produces eggs disomic for the X chromosome and eggs nullosomic for the X chromosome, while meiosis in the father is normal. The father's X- or Y-bearing sperm fertilize the three kinds of eggs (X, XX and 0) to produce the six types shown in the Punnett Square below. In contrast to Punnett Squares you have seen before, the frequency of the egg types shown as the column headings is not equal. The first column (normal meiosis to produce an X-bearing egg) is much more frequent than either of the exceptional types (XX and 0).

This produces normal males (XY), normal females (XX), inviable zygotes lacking the X chromosome (Y0), Klinefelter males (XXY), Turner females (X0), and a new type that we have not discussed: XXX. We will consider XXX individuals after examining the consequences of paternal nondisjunction.
Consider the case in which paternal nondisjunction produces five possible kinds of sperm:
These fertilize normal X-bearing eggs to produce the progeny types shown below.
In contrast to Down Syndrome, where most of the cases are the result of maternal nondisjunction, mainly at meiosis I, most females with Turner Syndrome have inherited their single X chromosome from their mother. This means that Turner Syndrome primarily results from paternal nondisjunction. Because the sperm resulting in a Turner Syndrome birth are nullosomic for the sex chromosomes, there is no information on the meiotic division at which the error occurred.
Most Turner Syndrome conceptions do not complete development. About 15% of spontaneous abortions are 45,X. Once individuals with Turner Syndrome are born, their survival is normal.
What are these new types, XXX and XYY? XXX individuals are phenotypically normal females who may never learn of their chromosomal abnormality. XYY individuals are phenotypically normal males who also may never learn of their chromosomal abnormality.
This leads us to ask whether the six karyotypes that we now know about with respect to sex chromosome aneuploidy can tell us anything about the role of sex chromosomes in development.
Because XX, X0, and XXX individuals are female, and XY, XXY and XYY are male, it appears that, in humans, the presence of one or more Y chromosomes causes maleness, and the number of X chromosomes is not a factor.
| Female | Male |
| XX | XY |
| X0 | XXY |
| XXX | XYY |
We leave as an exercise for the student to draw Punnett Squares for the matings shown below. Even without nondisjunction, there are some interesting results.
We turn our attention to Down syndrome, which is one of the few viable trisomies besides XXX. We can ask human genomics types for four numbers: the number of protein-coding genes in the genome, the number of such genes on the X chromosome, the Y chromosome, and chromosome 21. Different numbers are offered by different sources, it only matters that we get all four numbers from the same source at the same time. The results are shown in the figure below.

The X chromosome has more than three times the number of genes found on chromosome 21. Only the Y chromosome has fewer genes than chromosome 21. There are other chromosomes that have more genes than chromosome 21, but fewer than the X chromosome.
| Chromosome | Gene Count | Trisomy |
| X | 1376 | normal |
| 12 | 1370 | inviable |
| 14 | 1315 | inviable |
| 16 | 1098 | inviable |
| 15 | 998 | inviable |
| 20 | 768 | inviable |
| 22 | 749 | inviable |
| 13 | 545 | Patau syndrome very poor survival |
| 18 | 429 | Edwards syndrome poor survival |
| 21 | 360 | Down Syndrome good survival |
| Y | 284 | ? |
Down Syndrome (trisomy 21) is the autosomal trisomy with the greatest chance of survival. Trisomy for chromosome 18 or 13 has much greater effects than trisomy 21, and these types survive poorly. Six other chromosomes with fewer genes than the X chromosome produce inviable trisomics.
What is it about the X chromosome that permits XXX, XXXX, and XXXXX types to survive, while other chromosomes with fewer genes produce inviable trisomics? Is the X chromosome uniquely free of genes that produce phenotypes when present in extra copies, or is there another explanation?